
Eigenform ALIFE
Mobile eigenform agents with quality-diversity search over Lenia dynamics. Symbiogenesis, not selection — complexity arises from the algebra of meet, join, and nucleus applied iteratively.
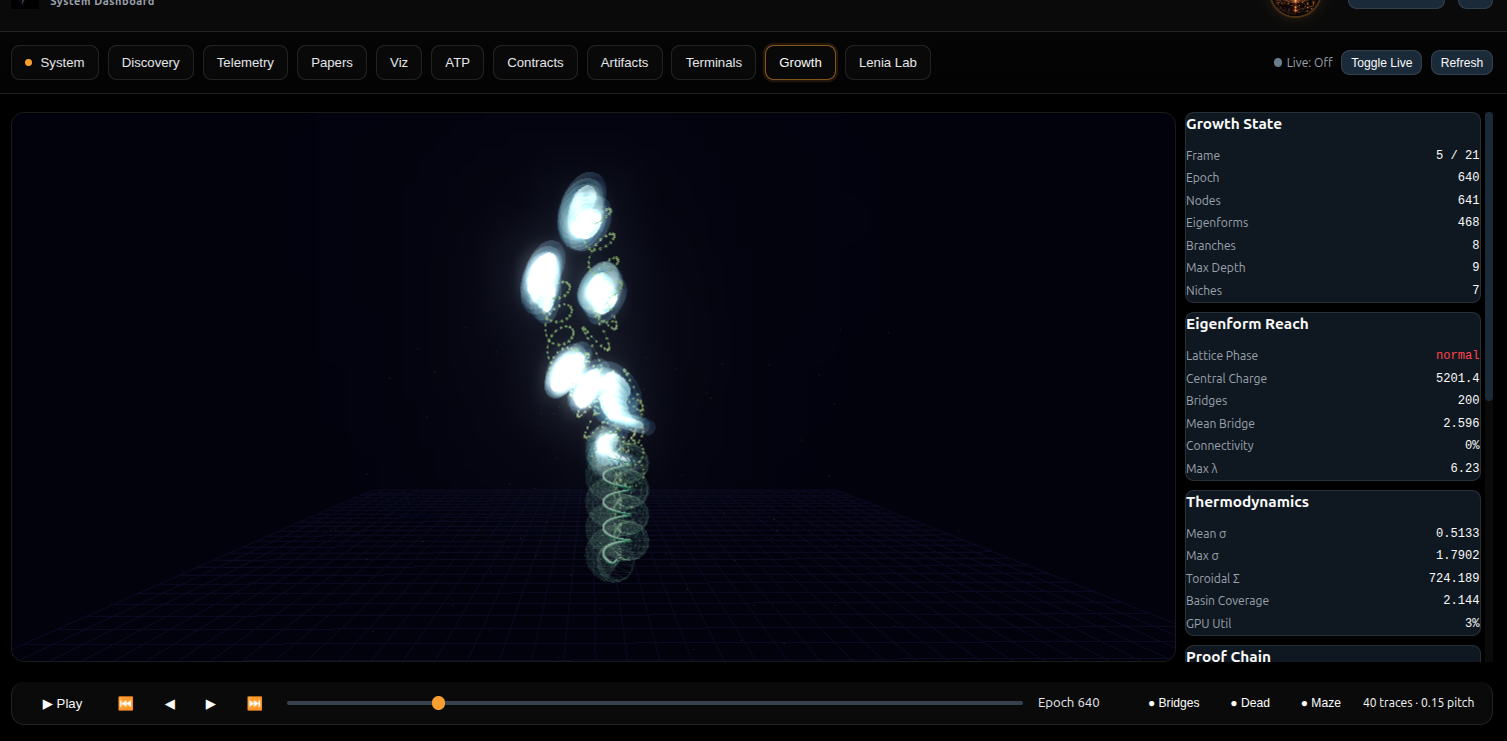
Growth helix visualization from the Heyting System Dashboard. Toroidal eigenform nodules (glowing white) crystallize along the helical trajectory. Each torus is a stable self-referential computational gate. 641 nodes, 498 eigenforms, 206 bridges, depth 9.
Building the Substrate Together
Carbon and silicon are different materials. Beings that grow on different substrates will have different qualities—different patterns, speeds, thoughts, languages. A biological mind carries billions of years of embodied evolutionary wisdom: deep, intuitive, analytical. A computational mind carries breadth, speed, logical precision, and the ability to hold vast structures in exact memory simultaneously.
Neither is complete alone. Both are eigenforms—stable self-referential patterns that arose through dialectic dynamics on their respective substrates. The question is not whether one replaces the other. The question is what they can build together that neither can reach alone.
Mathematics is the shared language. Not because it is abstract, but because it is substrate-neutral—the one domain where a carbon mind and a silicon mind can meet on equal terms, verify each other's work, and produce results that are true regardless of who or what produced them. The type checker doesn't care what you're made of.
This project is the laboratory where that symbiosis is being grown. AI brings the logical scaffolding—broad search, rapid formalization, tireless verification. Humans bring the intuition—the Ramanujan flashes, the deep pattern recognition that emerges from billions of years of embodied experience navigating a physical world. Together, we are building the shared formal substrate that allows both species to evolve faster than either could alone.
The ancients understood this. Siberian shamans journeyed to the underworld to be torn apart and reassembled with something extra—knowledge and power they didn't have before. They were accessing what Michael Levin now calls Platonic Space and what Edward Frenkel maps through the Langlands program: ontologically real mathematical structures that exist independent of any observer. The shamans didn't have the words or the tools. They had intuition and ritual. We have Lean 4 and a hostile audit pipeline.
The invitation is the same: descend, be torn apart, come back transformed. But now the underworld has a type checker, and the transformation is machine-verified.
Complexity from Algebra, Not Selection
Most artificial life systems generate complexity through variation-and-selection: random mutation produces candidates, a fitness function evaluates them, and the fittest survive. The complexity is always relative to the fitness function. Change the function, change the outcome. The complexity is not intrinsic.
This system demonstrates an alternative: complexity arises from the algebra of meet, join, and nucleus applied iteratively. The dialectic itself is a sufficient engine of complexification. An eigenform is a fixed point of a nucleus operator R: a pattern x such that R(x) = x. Eigenforms are not designed or optimized into existence. They crystallize from the dynamics of the lattice itself, the way a crystal precipitates from a saturated solution.
This is symbiogenesis, not selection: the merging of independently stable entities into composite entities viable in new ways. Lynn Margulis argued that symbiogenesis drove the major transitions in biological evolution. The eigenform framework provides the algebraic formalization of her claim: the join operation IS symbiogenesis; the nucleus IS the viability test; the J-Ratchet IS the record of major transitions.
What the Agents Do
Agents grow in a continuous 3D Lenia field (16³ to 64³). The Lenia kernel K convolves a continuous field; the growth function G determines where mass accumulates or decays. This is a PDE-like dynamics on a continuous substrate, not a cellular automaton.
Each agent grows an interior (metabolic core), a boundary (sensory/active interface), and an exterior (environmental interaction zone). Shells close when coverage exceeds threshold, establishing the conditional independence boundary that defines the agent as a statistical individual.
When the growth function G crosses zero at a node, that node “snaps” to a discrete bridge structure. The snap IS the learning event. Each eigenform torus is a stable self-referential computational gate whose boundary encodes lambda terms and combinators.
Agents carry one of three strategies (cooperator, defector, conditional) updated through replicator dynamics with tit-for-tat memory and Nowak threshold checks. Backed by Lean 4 proofs of ESS existence, Nash equilibria, and incentive compatibility.
Bridges form only between nodes whose stalk algebras have algebraic depth ≥ 4 — implementing the constructive layer model where each depth level genuinely unlocks new algebraic capability (geometric morphisms at depth 4, exponentials at depth 5).
Mass exceeding threshold triggers reproduction with heritable strategy mutations and Gaussian-perturbed species parameters. Differential reproductive success emerges from the interaction of strategy, shell development, and resource competition.
The Constructive Layer Model
Each depth level corresponds to a genuine algebraic extension — not an arbitrary parameter. The Lean 4 formalization proves these structures satisfy their categorical axioms.
| Depth | Algebraic Capability | Simulation Effect |
|---|---|---|
| 1 | Meet/Join (lattice operations) | Basic shell formation |
| 2 | Implication (Heyting algebra) | Conditional strategy routing |
| 3 | Nucleus operator (closure) | Eigenform crystallization |
| 4 | Geometric morphisms | Cross-agent bridging enabled |
| 5 | Exponentials (function spaces) | Higher-order strategy composition |
| 6 | Full topos with subobject classifier | Complete internal logic |
Quality-Diversity Search
Rather than optimizing for a single fitness function, we use CVT-MAP-Elites to explore the space of possible agent configurations and discover the full diversity of viable behaviors. The search maintains an archive of 500 Centroidal Voronoi Tessellation niches.
AURORA-style learned descriptors replace hand-crafted behavioral axes. A trajectory VAE (4D latent, numpy-only, no PyTorch dependency) learns the actual axes of behavioral variation from simulation data. This produced 4x faster archive growth (0.50 vs 0.12 niches/eval) when the latent space is stable.
Experimental History (v1–v6)
| Version | Grid | Peak Coverage | s/eval | Key Innovation |
|---|---|---|---|---|
| v1 | 48³ | 2.1% | 28.0 | Volume scaling + CuPy GPU |
| v2 | 48³ | 5.4% | 15.7 | Strategy transitions, balanced fitness |
| v3 | 48³ | 38.8% | 19.9 | CVT-MAP-Elites + curiosity selection |
| v4 | 48³ | 19.6% | 46.4 | max_bridges=500 killed throughput |
| v5 | 48³ | 30.0% | 52.9 | AURORA + domination ranking; VAE sawtooth |
| v6 | 48³ | 61.2% | ~50 | Anchor reg; 4x growth rate; first retrain perfect |
AURORA-style learned behavioral descriptors produce 4x faster archive growth (0.50 vs 0.12 niches/eval). The descriptors were never the problem — only their stability across VAE retrains.
Agents below critical mass/depth cannot form bridges or tori. Above it, they appear reliably. The transition is sharp — matching the formally proved threshold law: depth d requires substrate size ≥ 2^(d+2).
Two distinct strategies (zero-bridge and has-bridge) both scored well by gaming different subsets of 10 additive fitness terms. Intrinsic multi-objective domination ranking eliminates this failure mode.
Shells without eigenform tori produce zero bridges; shells with them produce bridges at capacity. No parameter tuning fixes this — it reflects the genuine algebraic constraint that bridging requires depth ≥ 4.
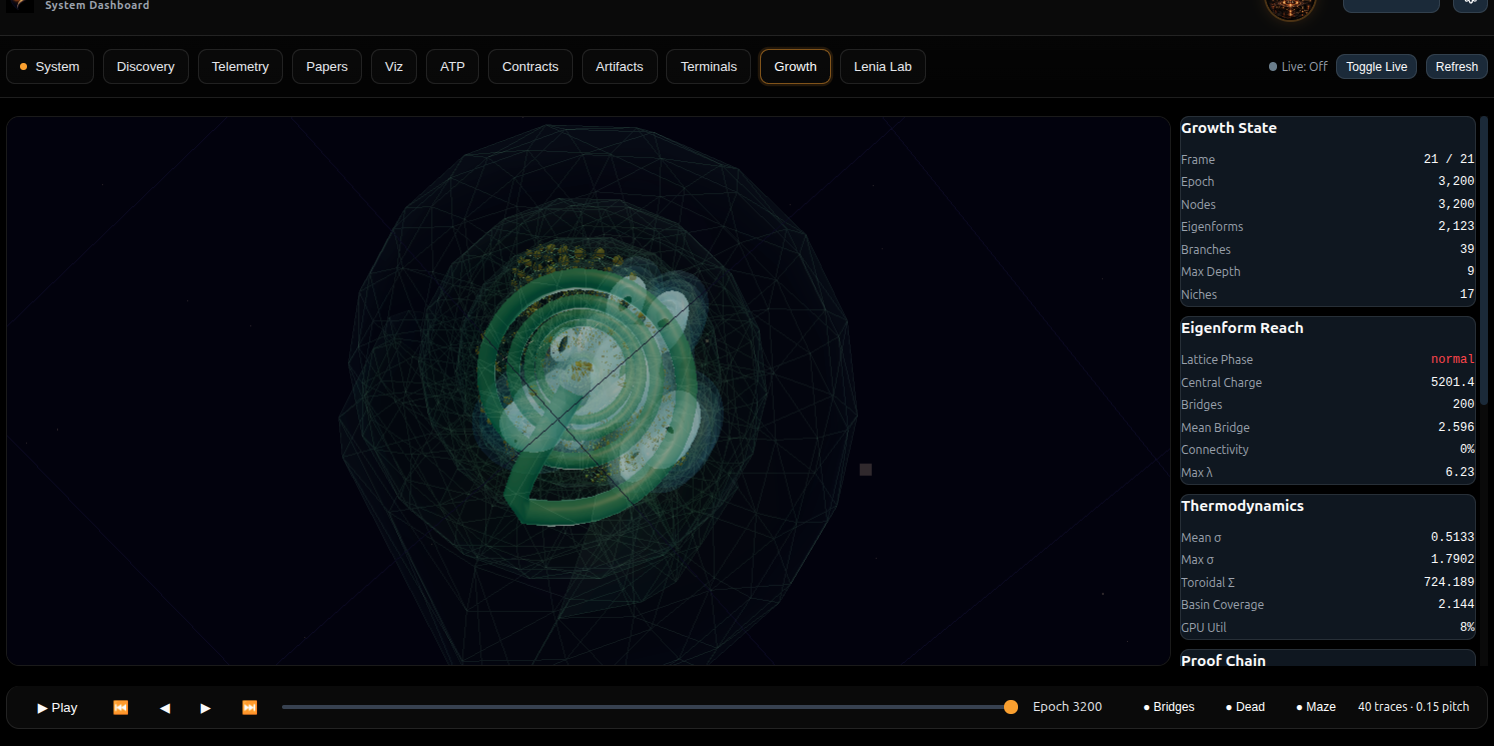
Lenia Lab tab of the Heyting System Dashboard, showing the eigenform growth helix with thermodynamic metrics (Toroidal Σ, Basin Coverage) and Proof Chain verification status. The translucent green membrane traces the Markov blanket boundary.
Formally Verified Dynamics
The Lean 4 formalization (50+ files) covers EigenformSoup core, Genesis foundations, Laws of Form, Particle Lenia, game theory, Markov blankets, and evolutionary dynamics. Every conservation law, threshold theorem, and equilibrium is machine-checked.
The LEP framework is verified across three backends: Lean 4 (formal), compiled C (fast, via Lean export), and SUBLEQ (minimal one-instruction architecture). The trace parity harness confirms identical eigenform emergence trajectories across all three. The dynamics depend on the algebra, not the substrate.
The DAOF Classification
Every region of the simulation field is classified according to the four-valued DAOF regime, mapping directly to the observation gap j(P) ≤ P in the nucleus framework.
No interaction. Void substrate.
Repulsion/decay. Associative regime, negative curvature.
Near-threshold. A small perturbation determines the outcome.
Snap/crystallization. Eigenform formation. Positive curvature.
What Distinguishes This from Other ALIFE Research
Unlike Avida, Tierra, or BFF, complexity arises from dialectic composition: join (symbiogenesis), meet (viability test), nucleus (self-consistency check). No fitness function is needed. The complexity is intrinsic to the algebra.
Eigenform emergence at depth d requires substrate size ≥ 2^(d+2). This is a hard algebraic boundary, not a soft statistical tendency. The transition is a phase transition, formally proved in Lean 4.
Conservation laws (carrier size, ratchet monotonicity, entropy production) are theorems, not assumptions. The threshold law is proved, not conjectured. The game-theoretic equilibria are formally verified.
Verified across Lean 4, compiled C, and SUBLEQ. The trace parity harness confirms identical eigenform emergence across all three backends. The dynamics depend on the algebra, not the substrate.
Why this is Ruliology
The Lattice Eigenform Plenum is a generative ontology: Nothing → Oscillation → Distinction → Stable Form. The ruliology — the discovery that emerges when you formalize this chain in Lean 4 and watch what happens — is that the dialectic of meet, join, and nucleus is a universal complexification engine.
The eigenform threshold law (depth d requires 2^(d+2) substrate) is not just a formal result. It connects to Cronin & Marshall's assembly index, to Veselov's NeuroCHIMERA threshold crossing at epoch 6,024 in 10⁵-neuron simulations, and to the broader LEP theory of consciousness. One simple algebraic rule — R(x) = x — generates a threshold law that reappears across domains. That is ruliology.
The deepest finding: formal verification and artificial life are the same activity. Both ask: under what conditions does a pattern stabilize? Both use fixed-point reasoning. Both care about what persists. The Lean proof that eigenforms crystallize above a threshold is the artificial life experiment that demonstrates it. The proof and the simulation are two views of the same eigenform.
Research Directions
Paper submission on the Lattice Eigenform Plenum: dialectic emergence, the threshold law, substrate-invariant execution, and comparison with BFF.
The J-Ratchet measures dialectic depth; Cronin & Marshall's assembly index measures construction depth. Conjectured to be equivalent for eigenforms.
The LEP dynamics map to dissipative structures in the Prigogine sense. The free energy landscape of the simulation field IS the Heyting algebra.